





Россия
Россия
УДК 626.872 Мелиорация низинных болот
УДК 35.073.515.11 Амелиорация
УДК 631.6 Сельскохозяйственная мелиорация
ГРНТИ 68.31 Сельскохозяйственная мелиорация
ОКСО 35.00.00 Сельское, лесное и рыбное хозяйство
ББК 4 СЕЛЬСКОЕ И ЛЕСНОЕ ХОЗЯЙСТВО. СЕЛЬСКОХОЗЯЙСТВЕННЫЕ И ЛЕСОХОЗЯЙСТВЕННЫЕ НАУКИ
ТБК 5607 Сельскохозяйственная мелиорация
BISAC TEC003000 Agriculture / General
В статье рассматривается необходимость использования уникальной способности ризосферных микроорганизмов фиксировать атмосферный азот и являться источником биологического азота для сельскохозяйственных растений. Проанализированы имеющиеся данные о механизмах ассоциативной азотфиксации, масштабах и значение этого процесса при микробно-растительных взаимоотношениях. Детально изучены и систематизированы факторы, оказывающие существенное влияние на процесс азотфиксации и определяющие активность деятельности ассоциативных диазотрофов, эффективность всего процесса азотфиксации. Представлена функциональная схема растительно-микробных взаимодействий в почве, раскрывающая основные механизмы этого процесса, а также схема комплекса математических моделей микробиологических процессов для описания биогеохимической трансформации, накопления или расходования веществ в почве. Исследования закономерностей и особенностей взаимодействия диазотрофов с растениями, моделирование и анализ изменений количественных показателей их развития под влиянием различных факторов, включение алгоритмов этих моделей в комплексные модели продуктивности агроэкосистем позволят более грамотно и обоснованно подходить к планированию мелиоративных режимов, разработке агробиологических технологий и регулированию почвенного плодородия на мелиорируемых землях.
диффузный сток, поглотительные колодцы, дренажный сток, пруды-накопители, коллекторы–накопители, точное орошение, очистка дренажного стока, водооборотные системы
Введение. В современных условиях развития сельского хозяйства России при широком использовании адаптивно-ландшафтных систем земледелия неизменно растет интерес к дополнительным источникам элементов питания растений, особенно азотного [1]. Величина урожаев сельскохозяйственных культур, наряду с другими лимитирующими факторами, в значительной степени определяется обеспеченностью растений азотом. Основные запасы азота (75…80%) сосредоточены в атмосфере в молекулярном виде и недоступны растениям. Наибольшая часть природного азота вовлекается в круговорот за счет симбиотической и несимбиотической фиксации в процессе жизнедеятельности азотфиксирующих микроорганизмов, обеспечивая в значительной мере жизнь на нашей планете. Масштаб биологической фиксации азота на земной суше составляет до 200 млн т в год (для сравнения: за счет химической промышленности в мире производится всего 84 млн т азотных удобрений). Сельскохозяйственные растения получают 2/3 необходимого им азота из азотного резерва почв, созданного и поддерживаемого деятельностью микроорганизмов-диазотрофов [2]. Вклад биологической азотфиксации в сельское хозяйство достаточно высок, примерно вдвое превосходит вклад химических азотных удобрений, а в ежегодном потоке азота на земной суше почти в 3 раза больше, чем вклад азота минеральных удобрений [1].
Недостаток азота минеральных удобрений и необходимость увеличения продуктивности сельскохозяйственных культур при стремлении снизить энергетические затраты на производство продукции растениеводства требуют комплексного применения минерального и биологического азота. Еще в середине ХХ века Д.Н. Прянишников обращал внимание на то, что построение бездефицитного баланса в земледелии должно состоять в сочетании двух путей: 1) первоочередное обеспечение техническим азотом наиболее ценных культур; 2) покрытие недостающей потребности сельскохозяйственных культур «за счет увеличения других источников прихода азота, главным образом азота биологического» [3].
В первой половине 20 в. наблюдался неуклонный рост научно-исследовательских работ по исследованию микробиологических процессов и созданию перспективных микробных препаратов для различных культур. Однако после второй мировой войны бурное развитие химической промышленности и решение вопросов технологии выращивания сельскохозяйственных культур с использованием химических удобрений отодвинули эти исследования на второй план. Но в последние годы в связи с возникновением проблемы экологизации сельскохозяйственного производства наступил период переосмысления методов и стратегии использования биологических и химических источников азотного питания сельскохозяйственных растений [4], поиска новых микробно-растительных азотфиксирующих сообществ, разработки комплекса агробиологических технологий с целью повышения продуктивности азотфиксации в растениеводстве, роль которых и практическая значимость до настоящего времени недооценивается.
Учитывая глобальное значение биологической фиксации в круговороте азота в биосфере (до 70…90% азота в пахотных почвах фиксируется из атмосферного воздуха симбиотическими, ассоциативными и свободноживущими микроорганизмами [5]), а также уникальность биохимических и физико-химических процессов фиксации азота микроорганизмами, внимание специалистов биологов, биохимиков и представителей сельскохозяйственных наук должна быть сфокусированы на всестороннем и глубоком исследовании явления азотфиксации, механизмов взаимоотношения растений и азотфиксирующих микроорганизмов, в том числе с использованием методов математического моделирования.
Материалы и методы.
Целью мелиорации сельскохозяйственных земель является расширенное воспроизводство плодородия почвы, получение рациональной урожайности сельскохозяйственных культур при экономном расходовании всех видов вовлеченных ресурсов и недопущении или компенсации ущерба окружающей среде.
Сущность мелиорации сельскохозяйственных земель заключается в качественном целенаправленном изменении состава и свойств почв и управлении почвенными процессами. Традиционно при проведении мероприятий по улучшению земель уделяется доминирующее внимание гидромелиорации, совершенно справедливо учитывая ее комплексное воздействие на почвы, ландшафты, природу. Совершенно очевидно, что эффектность мелиоративного воздействия зависит от точного регулирования мелиоративного режима почв, земель, агроландшафтов, что становится возможным при создании независимых от естественных причин условий, для максимально возможного проявления пищевого, энергетического и водного факторов существования, трансформации и развития, сохранения агроценозов, обладающих нужными для человека свойствами. В последние годы в мелиоративном научном сообществе уделяется внимание развитию направлений, связанных с изучением биотической составляющей протекающих в почве процессов, которые фактически и определяют важнейшее свойство почвы – плодородие. Почвенная биота (растения, грибы, бактерии, водоросли, микроорганизмы, простейшие. инфузории, жгутиковые, членистоногие насекомые, черви, моллюски и другие) участвует во множестве биогеохимических процессов переноса, трансформации, минерализации, гумификации, накопления, перераспределения биогенных элементов и веществ в почве, структурируя и формируя ее важнейшие свойства. Настоящая статья посвящена вопросам ассоциативной биотической фиксации азота живущими в почве бактериями диазатрофами и разработке подходов к установлению значений показателей мелиоративного режима, обеспечивающих накопление этого биогена, обеспечивающего высокий уровень плодородия почвы.
Концептуальной базой статьи является принципиально новый научный подход к обоснованию методов мелиорации и рекультивации земель с учетом современных представлений о совокупности процессов при мелиорации, протекающих в агроландшафтах. В частности, механизмов взаимодействия растений и азотфиксирующих микроорганизмов диазотрофов, физических и физико-химико-биологических основ явления азотфиксации. По нашему мнению, это также необходимо для поиска новых микробно-растительных азотфиксирующих сообществ, разработки комплекса агробиологических технологий для повышения продуктивности азотфиксации в растениеводстве и для повышения продуктивности сельскохозяйственных земель.
Для изучения основных аспектов взаимодействия ассоциативных диазотрофов с сельскохозяйственными растениями при осуществлении мелиоративной деятельности были применены информационно-аналитические методы исследования, которые включали комплексный анализ, структуризацию научной информации.
Результаты и обсуждение. Известно, что 70…75% азота в составе биомассы урожая сельскохозяйственных культур имеет биотический источник своего происхождения (это азот, фиксированный диазотрофами, и азот минерализованной части органического вещества почвы также преимущественно микробиологического происхождения). В связи с этим азотфиксация выделена, наряду с фотосинтезом, в ряд основных физиологических процессов, а биологический азот рассматривается как фактор сохранения плодородия почвы и формирования продуктивности сельскохозяйственных культур [1].
В наибольших масштабах азот фиксируется в ходе ассоциативной фиксации, при взаимодействии бактерий и растений, не образующих специализированных органов (клубеньков) на корнях и стеблях. Ассоциативные азотфиксирующие бактерии широко распространены в почвах различных климатических зон, что дает возможность их широкого использования в сельском хозяйстве. Установлена высокая отзывчивость ряда сельскохозяйственных культур на обработку их биопрепаратами азотфиксаторов [3].
Биологический азот имеет и важное экологическое значение, так как поступает в почву постепенно, в течение всей вегетации, что исключает избыточное его накопление и снижает опасность загрязнения растениеводческой продукции и окружающей среды. Экологической составляющей азотфиксации является и то, что азот, получаемый в результате микробной фиксации из атмосферного воздуха, не может загрязнять окружающую среду (почвы, водоемы), поскольку поступает в метаболические пути и используется для биосинтеза азотсодержащих соединений [1]. Преимущество биологического азота не только в безвредности. Для его накопления требуются относительно небольшие затраты энергии на активацию азотфиксирующих микроорганизмов. При биологической фиксации источником энергии является, как правило, продукты фотосинтеза, фиксированный азот усваивается растениями практически полностью [5].
Населяющие ризосферу и ризоплану бактерии образуют с корневой системой растений прочные ассоциации и формируют специфические ризосферные бактериальные сообщества, образуя многокомпонентную интегральную систему с новыми свойствами, детерминированными взаимодействием партнеров.
Установлено, что около 30…50% продуктов фотосинтеза растения оставляют в ризосфере в виде корневых экссудатов и ризодепозитов. Корневые выделения и корневой опад растений являются энергетическим субстратом для ассоциативных бактерий-диазотрофов. Повышение азотфиксирующей активности микроорганизмов в ризосфере растений обеспечивает депонирование доступного для растений азота в большинстве природных экосистем. Уже к середине 80-х годов было продемонстрировано широкое распространение ассоциативной азотфиксации в ризосфере растений различных семейств и в различных почвенно-климатических условиях: этот процесс протекает практически во всех почвах в ризосфере растений самых разных мест обитания и осуществляется многими представителями родов Azospirillum, Azotobacter, Agrobacterium, Acetobacter, Klebsiella, Enterobacter, Flavobacterium, Pseudomonas, Bacillus, Clostridium и др. [6]. Вышеперечисленные бактерии характеризуются наличием одинакового ферментного комплекса – нитрогеназы, который выступает катализатором процесса азотфиксации [1, 3].
Микроорганизмы, усваивающие молекулярный азот, или диазотрофы имеют сходный биохимический механизм фиксации молекулярного азота воздуха, в основе которого лежит процесс восстановления N2 по уравнению: N2 + 6е- + 6Н+ → 2NН3.
При использовании ассоциативных диазотрофов в зависимости от почвенно-климатических условий в агроценозы вовлекается 30…60 кг/га азота из воздуха [1]. По другим источникам [6] масштабы ассоциативной азотфиксации в зоне умеренного климата достигают 50…150 кг/га молекулярного азота за вегетационный период, в тропических широтах – 170…600 кг азота на гектар в год. Кроме того, интродуцируемые микроорганизмы, применяемые для инокуляции семян сельскохозяйственных культур, стимулируют рост и развитие растений, повышают их устойчивость к биотическим и абиотическим факторам, в том числе ограничивают рост фитопатогенов, регулируют поступление загрязнителей из окружающей среды в продукцию сельскохозяйственного производства [5]. По обобщенным данным разных исследований применение биопрепаратов, созданных на основе штаммов ассоциативных микроорганизмов, приводит к росту урожайности зерновых в среднем на 15…20%, овощных и других сельскохозяйственных культур – на 20…30% [1].
Использование во второй половине XX века высокочувствительных и точных методов регистрации активности азотфиксации в почве (изотопный и ацетиленовый методы) позволило выявить новые группы микроорганизмов, обитающих в ризосфере растений и участвующих в этом процессе, а также дала возможность найти решение в управлении активностью ассоциации растение–азотфиксирующие бактерии, что открыло большие возможности для применения ассоциативной азотфиксации в сельском хозяйстве [3]. Установлено, что нитрогеназная активность выявлена у более 50% бактерий, выделенных из ризосферы небобовых растений, обнаружена повышенная активность азотфиксации в фитоплане (ризосфере и филлосфере) небобовых растений. Среди сельскохозяйственных культур наиболее изучена ассоциативная фиксация азота в ризоплане сахарного тростника, пшеницы, ржи, ячменя, кукурузы, сорго, овсяницы луговой, ежи сборной, овощных культур. Установлена высокая отзывчивость ряда сельскохозяйственных культур (особенно злаковые растения) на обработку биопрепаратами азотфиксаторов. Положительное действие на рост и продуктивность растений установлено для многих штаммов бактерий, относящихся к разным систематическим группам, причем механизм этого действия не всегда известен [1, 3].
Ассоциативная азотфиксация проходит в ризосфере небобовых растений во всех типах почв, но с различной скоростью. В исследованиях количественные параметры несимбиотической азотфиксации постоянно подвергались существенной переоценке. По результатам исследований третьей четверти ХХ века размеры несимбиотической азотфиксации под различными культурами (зерновыми, коноплей, хлопчатником) и на разных типах почв (дерново-подзолистых, серых лесных, черноземных почвах и сероземах) составили 30…40 азота на гектар в год. В дерново-подзолистых почвах при наличии активно вегетирующей растительности (ячменя, злаковых трав) суммарная продуктивная ассоциативная и несимбиотическая азотфиксация оценивается на уровне 40…55 кг азота на гектар, в парующей почве – 10…13 кг азота на гектар в год. В почве рисовых полей за счет несимбиотической фиксации атмосферного азота ежегодно накапливается до 60…70 кг азота на гектар. В посевах яровой пшеницы за счет ассоциативной азотфиксации ежегодное поступление азота достигает 20…30 кг азота на гектар [1].
Видно, что поступление атмосферного азота при формировании биомассы сельскохозяйственных растений за счет ассоциативной азотфиксации значительно варьирует на разных почвах и в различных климатических условиях. Очевидно также, что поступление ассоциативного азота в конкретный агроценоз определяется видом возделываемой культуры и условиями ее выращивания, которые оказывают существенное влияние как на состав диазотрофов, так и на их азотфиксирующую активность [3]. Действие растений на ассоциативную фиксацию азота заключается в обеспеченности последних продуктами экзосмоса и корневого опада: до 1/3 всего углерода фотосинтеза прижизненно выделяется растениями в почву в виде богатых энергией, легкодоступных для микроорганизмов углеводов, стимулирующих азотфиксацию вне зависимости от свойств почв. Количество прижизненного легкодоступного для диазотрофов органического опада и выделений растений, поступающих в прикорневую зону, варьирует в пределах 10…30% всех продуктов фотосинтеза [1,3].
Факторы, определяющие эффективность ассоциативной азотфиксации и особенности функционирования системы растение – ассоциативные диазотрофы можно условно разделить на энергетический фактор, водный фактор и пищевой фактор.
Фиксация молекулярного азота ассоциативными микроорганизмами осуществляется при постоянном притоке энергии и наличии источников электронов, необходимых для функционирования нитрогеназного комплекса. В посевах уровень азотфиксации в почве значительно выше, чем при отсутствии посевов, поскольку корневые выделения и корневой опад растений являются энергетическим субстратом для ассоциативных бактерий-диазотрофов. Кроме того, в ризоплане создаются благоприятные условия для поддержания высокой активности нитрогеназы, корни растений быстро поглощают азотсодержащие метаболиты азотфиксаторов, а муцигель защищает нитрогеназу от избытка кислорода. Изменения интенсивности фотосинтеза в течение онтогенеза растений является причиной изменений величины азотфиксации на протяжении вегетационного периода [3, 6].
Важным условием, эффективной ассоциативной азотфиксации, является применение биологических препаратов, созданных на основе активных штаммов микроорганизмов, обладающих повышенной способностью к ассоциации с культурными растениями и интенсивной азотфиксацией. Инокуляция сельскохозяйственных растений микроорганизмами позволяет повысить продуктивность посевов во всех регионах РФ (однако при недостатке увлажнения в период вегетации урожайность от инокуляции не изменяется). Применение ряда препаратов в благоприятные по погодным условиям годы обеспечивает получение урожайности злаковых культур в таких же размерах, как внесение азотного удобрения из расчета 30…45 кг азота на гектар. [7]. При количестве осадков, близком к среднемноголетней норме, применение ассоциативных биопрепаратов эквивалентно внесению азотного удобрения под озимые пшеницу, рожь и тритикале, а также ячмень и овес из расчета 30 кг азота на гектар, под яровую пшеницу – 30…45 кг азота на гектар, под кукурузу – 45…60 кг азота на гектар и под картофель 40…45 кг азота на гектар [1, 3]. Положительные результаты от инокуляции препаратами Azospirillum, выраженные прибавкой урожая на 5…40%, получены зарубежными исследователями. Наибольшую прибавку урожайности зерна яровой пшеницы и ячменя от инокуляции биопрепаратами ассоциативных диазотрофов (3 и 2,2 ц/га соответственно) на фоне РK-удобрений можно получить при содержании гумуса в почве 2,5…2,7%, нейтральной реакции почвенной среды (рНKCl 6,5…6,7 и 6,1…6,7 соответственно) и оптимальном увлажнении (ГТК за май = 1,0…1,3 и ГТК за май–июнь = 1,3…1,8). При недостатке влаги (ГТК за май < 0,8) или повышенном увлажнении (ГТК за май > 1,8) эффективность препаратов на основе ризосферных ассоциативных диазотрофов снижается [1]. Установлено, что внесение в почву активных штаммов ризосферных микроорганизмов, несмотря на большую вариабельность показателей, в подавляющем большинстве случаев обеспечивает существенный рост интенсивности связывания атмосферного азота [8].
Активность ассоциативных диазотрофов и в целом процесса азотфиксации определяются естественными природными условиями или теми условиями, которые обеспечиваются мелиоративными приемами:
- климат, температурный режим и свойства атмосферного воздуха;
- тип почвы, воднофизические и биохимические свойства почвы;
- содержание, качественный состав и распределение органического вещества по профилю почвы;
- наличие и содержание в почве подвижных соединений азота, фосфора и калия;
- реакция почвенной среды;
- численность, состав и взаимодействие микробных сообществ;
- влажность почвы, интенсивность выпадения осадков, положение уровня грунтовых вод, химические и электролитические свойства осадков, почвенной влаги и грунтовых вод.
На рисунке 1 представлена функциональная схема растительно-микробно-диазотрофного взаимодействия в почве.
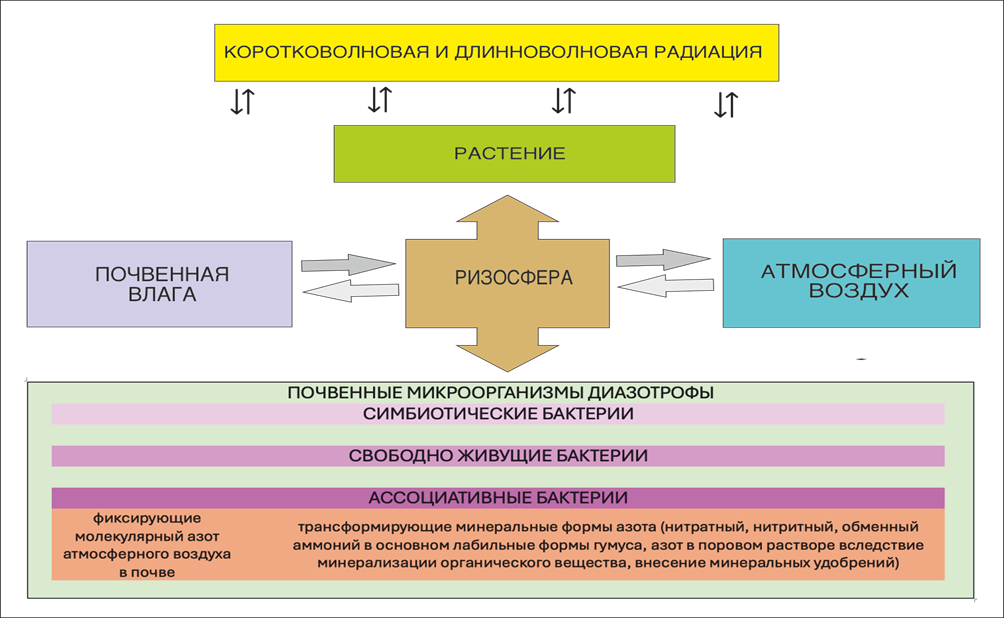
Рис. 1. Схема растительно-микробно-диазотрофного взаимодействия в почве.
Уровень активности фиксирования атмосферного азота диазотрофами снижается в ряду черноземы – серые лесные – дерново-подзолистые почвы, что обусловлено структурой микробиоценоза (21…26 – видов диазотрофов в черноземах, около 15 – в серых лесных почвах, 2…4 вида диазотрофов в подзолистых почвах) [8]. По мере увеличения влажности почвы интенсивность связывания атмосферного азота диазотрофами возрастает, при полной водонасыщенности (вследствие развития негативных анаэробных процессов) – снижается. Потенциальная активность азотфиксации больше на известкованных почвах, границы реакции почвенного раствора, в которых ассоциативные диазотрофы имеют наиболее благоприятные условия для развития, находится в пределах рНKCl 5,8…7,0. Температура верхнего слоя 0…10 см почвы в течение вегетации слабо влияет на азотфиксацию, однако существует большая зависимость ассоциативной азотфиксации в филлосфере от изменений температуры воздуха, оптимальная величина которой находится в пределах 20…34°С [1].
Органические вещества, образующиеся при разложении растительных остатков, также в значительной мере регулируют состав и численность почвенной микрофлоры, в том числе диазотрофов [6]. Активизация азотфиксации происходит и при внесении в почву соломы и других растительных остатков. Она более активна при соотношении углерода к азоту в растительных остатках и имеет большие значения при более узком соотношении C/N [1].
Интересно, что растения и микроорганизмы легко усваивают минеральный азот, но с повышением его содержания в почве активность ассоциативной азотфиксации, как правило, снижается. Распространено мнение, что минеральный азот в почве тормозит процесс ассоциативной азотфиксации, вместе с тем по другим данным, только высокие дозы азотных удобрений (100…150 кг/га) могут кратковременно подавлять ассоциативную азотфиксацию, а на фоне применения «стартовых» доз азотных удобрений азотфиксация на 30…45% больше, чем без них. Внесение азота в дозах, не превышающих физиологическую потребность растений (30…80 кг/га в зависимости от культуры), усиливает азотфиксацию и участие биологического азота в питании растений [1, 3].
Несмотря на значительные успехи в исследованиях процессов накопления, выноса, трансформации различных форм азота в почве, ассоциативной азотфиксации в агроценозах,
до настоящего времени отсутствуют надежные количественные оценки размеров поступления фиксированного азота почвы в конкретных условиях [9], поэтому разработка современных методов количественной оценки этих процессов, основанных на многофакторных математических биогеохимических моделях, является актуальной. Такие модели необходимы для познания потенциала взаимовыгодной интеграции растений и микроорганизмов, происходящей в ризосфере, в зависимости от скорости выделения углеродного субстрата корнями растений и конкурентных взаимоотношений с нефиксирующей азот частью ризосферной микрофлоры, определения степени влияния на эти взаимодействия микробиологического, растительного и почвенного факторов.
На рис. 2. представлен комплекс математических моделей микробиологических процессов для описания биогеохимической трансформации, накопления или расходования веществ в почве.
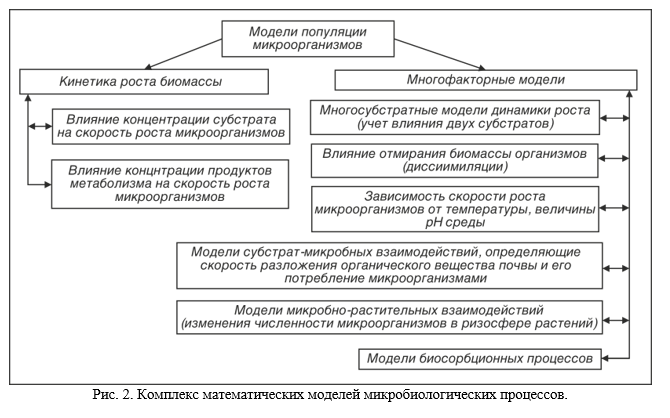
Характер интеграционных процессов в микробном ценозе ризосферы является важным показателем степени и направленности изменения почвенных условий при возделывании различных сельскохозяйственных культур, а, следовательно, может служить основанием для разработки путей их регулирования. Разработка такого рода моделей создает основы для регулирования роста ризобактерий при широком варьировании различных кинетических параметров роста микроорганизмов и почвенных условий, оценки поведения интродуцируемых штаммов ассоциативных азотфиксаторов в почвенных условиях в ризосфере различных растений и в итоге могут использоваться при разработке предложений для конструирования высокопродуктивных растительно-микробных сообществ, поскольку до настоящего времени при создании современных технологий в земледелии не учитываются пути использования почвенной микрофлорой и способы влияния на их эффективность дополнительного углерода, непрерывно выделяемого во внешнюю среду растениями в процессе своей жизнедеятельности (более 30% от всего углерода, ассимилированного растением).
Из множества необходимых для жизнедеятельности пищевых компонентов, субстратов, потребляемых организмами исследуемой популяции, субстрат, находящийся в наименьшей доступности и лимитирующий ростовые процессы, определяет скорость размножения. Этот принцип известен как принцип минимума Либиха.
Основное уравнение роста популяции представляет собой дифференциальное уравнение первого порядка с разделяющимися переменными, которое можно записать в виде:
![]()
микроорганизмов, ч –1.
Решением этого уравнения является выражение, позволяющее рассчитывать накопление биомассы в экспоненциальной фазе роста:
 .
.
Для полноценного использования и подбора моделей роста популяции азотфиксирующих бактерий в почве необходима полноценная параметризация, основанная на проведении натурных исследований в различных условиях для конкретных агробиоценозов.
При моделировании динамики роста микробиологического сообщества азотфикирующих бактерий в почве используется принцип лимитирующего фактора роста. При этом, принимается следующая логика: если при небольшом увеличении фактора наблюдается ответное увеличение удельной скорости роста, то данный фактор лимитирует рост; в противном случае фактор не является лимитирующим. Рост может быть лимитирован по одному или нескольким компонентам питательной среды, соответственно, говорят об однофакторном и многофакторном лимитировании.
Рост, лимитированный по концентрации одного из субстратов, описывается уравнением Моно:
![]()
![]()
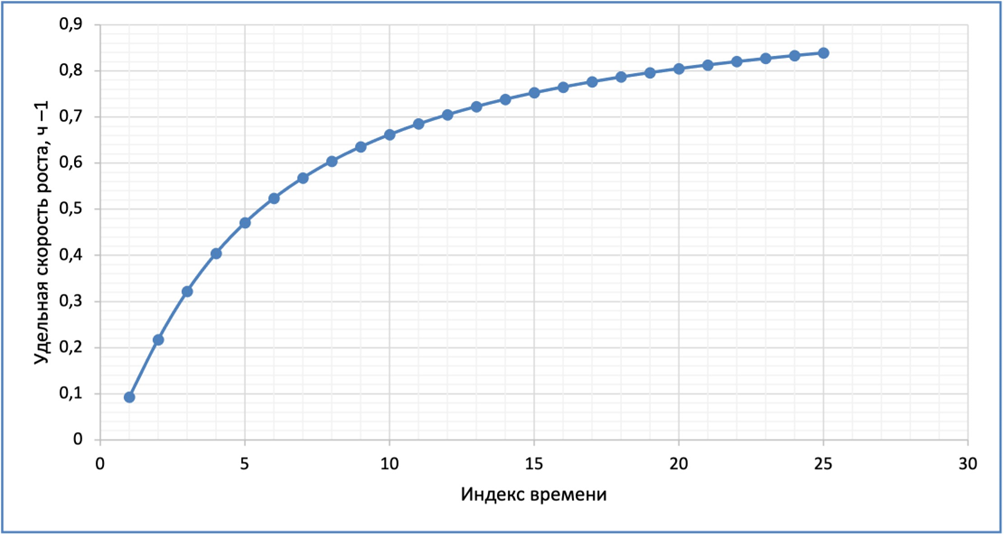
Рис. 3. Удельная скорость роста популяции азотфиксирующих бактерий в почве.
Константы Моно для бактерий имеют очень небольшие значения. Для гетеротрофов типичные значения константы Моно по органическому субстрату, =1…10 мг/л, а по кислороду (для аэробов)  =0,1…0,2 мг/л. Для бактерий диазотрофов, относящихся к автотрофам,
=0,1…0,2 мг/л. Для бактерий диазотрофов, относящихся к автотрофам,  =1 мг/л. (в расчетах, значения констант приняты по данным из открытых источников информации).
=1 мг/л. (в расчетах, значения констант приняты по данным из открытых источников информации).
Удельная скорость роста азотфиксирующих бактерий (для многофакторного лимитирующего роста бактерий, при увеличении в поровом растворе почвы концентрации амонийного азота от 0,48 до 12 мг/л и кислорода от 0.,08 до 2,0 мг/л), ч –1.
Выводы
- Исследование таких аспектов жизнедеятельности почвенных микроорганизмов, как биологическая фиксация азота и взаимоотношения с высшими растениями, в том числе с использованием методов математического моделирования, будут способствовать более грамотному и обоснованному планированию мелиоративных режимов, регулированию почвенного плодородия и продуктивности сельскохозяйственных культур.
- Модели позволяют концентрировать и развивать фундаментальные знания о изучаемых процессах, осуществлять количественную оценку и прогнозировать развитие процессов во времени, разрабатывать на их основе новых микробиологических препаратов, методов и дозировки их внесения в почву, установления требуемых значений показателей мелиоративного режима.
1. Завалин А.А. Ассоциативная азотфиксация и практика применения биопрепаратов в по-севах сельскохозяйственных культур / А. А. Завалин, А. А. Алферов, Л. С. Чернова // АГРОХИМИЯ, 2019, № 8, с. 83–96.
2. Умаров М.М. Азотфиксация в ассоциациях организмов // Проблемы агрохимии и эколо-гии, 2009. № 2. – С. 22–26.
3. Алферов А.А. Ассоциативный азот, урожай и устойчивость агроэкосистемы. – М.: РАН, 2020. – 184 с.
4. Игнатов В.В. Биологическая фиксация азота и азотфиксаторы // Соросовский образова-тельный журнал, № 9, 1998. – С. 28-33.
5. Тихонович И.А. Перспективы использования азотфиксирующих и фитостимулирующих микроорганизмов для повышения эффективности агропромышленного комплекса и улучшения агроэкологической ситуации РФ / И.А. Тихонович, А.А. Завалин // Плодоро-дие, 2016.– № 5. – С. 28–32.
6. Феоктистова Н.В., Марданова А.М., Хадиева Г.Ф., Шарипова М.Р. Ризосферные бакте-рии // Учен. зап. Казан. ун-та. Сер. Естеств. науки. – 2016. – Т. 158, кн. 2. – С. 207–224.
7. Карашаева А. С. Ассоциативные диазотрофы и их взаимодействие с растениями // Моло-дой ученый. 2016. № 6 (110). – С. 350-353. – URL: https://moluch.ru/archive/110/26656/ (дата обращения: 20.11.2023).
8. Шотт П.Р. Биологическая фиксация азота в однолетних агроценозах лесостепной зоны Западной Сибири: Автореф. дисс. на соискание уч. степени доктора сельскохозяйствен-ных наук. – Барнаул, 2007. – 39 с.
9. Кравченко Л.В. Роль корневых экзометаболитов в интеграции микроорганизмов с расте-ниями: Автореф. дисс. на соискание уч. степени доктора биол. наук. – М., 2000. – 51 с.